Virtually all life on earth relies on photosynthesis, a process carried out mostly by green plants using light in the blue and red regions of the visible light spectrum to generate reductant (ferredoxin and NADPH) and ATP used to power the synthesis of carbohydrates from carbon dioxide (CO2). This process splits water into protons, electrons and oxygen. The protons and electrons are used to synthesize ATP and NADPH, and the oxygen evolved is liberated mostly to the atmosphere.
This process was "invented" by cyanobacteria probably around 3 billion years ago, and is responsible for the great oxygenation event in earth's history, transforming the planet from an anaerobic to an aerobic environment. This process has led to today's 21% atmospheric oxygen levels that allow all animals to generate energy from mitochondrial respiration of plant-derived carbohydrates.
It is estimated that about 1.2 billion years ago a primitive eukaryotic organism containing a mitochondrion and nucleus engulfed a cyanobacterium but instead of digesting it, the organism retained it. This single primary endosymbiosis event likely gave rise to the entire green plant lineage with the cyanobacterium becoming the green chloroplast typical of the vast majority of plants that we know today. The initial photosynthetic eukaryote derived from this endosymbiosis event was a unicellular green algae. Plants did not colonize land until about 500 million years ago, slowly evolving into mosses, ferns, gymnosperms, cycads and ultimately angiosperms (flowering plants) about 200 to 140 million years ago (however, the exact time is still under considerable debate).
The chloroplast has a double outer membrane, its own DNA (resembling that of cyanobacteria) and its own DNA replication, transcription, translation and protein synthesis machinery. However there has been significant exchange of DNA between the nucleus and chloroplast during the last 1.2 billion years. Chloroplasts are maternally inherited in plants.
Photosynthesis takes place in the cyanobacterium-derived organelles called chloroplasts. The chloroplasts have inner stacks of lipid bilayer membranes (termed thylakoids) with a series of protein complexes and pigments embedded within the membranes that catalyze the light reactions of photosynthesis.
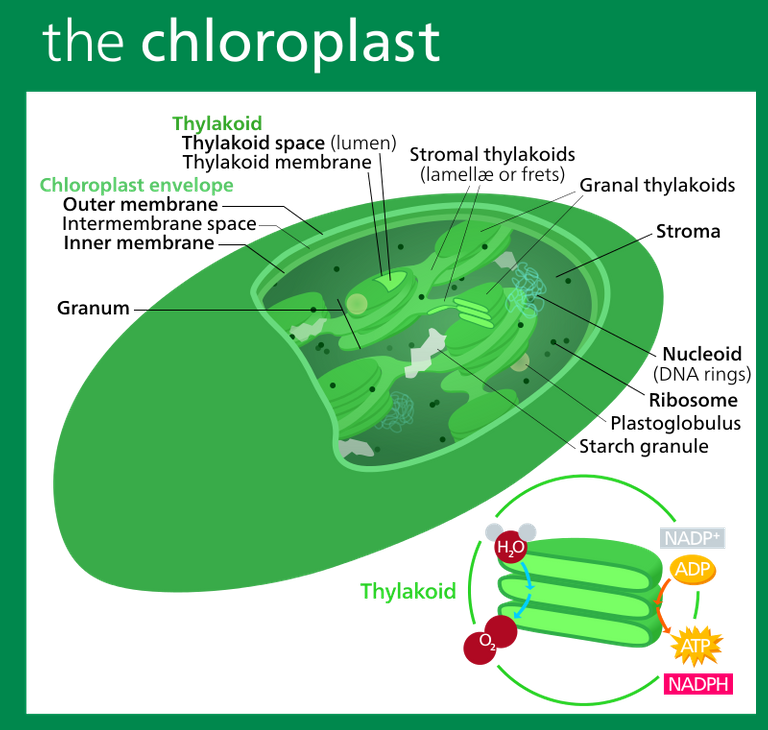
Structure of a typical higher plant chloroplast. Image Source: Chloroplast - Wikipedia
The thylakoid stacks somewhat resemble parking garage structures with several levels (granal stacks), connected by tunnels (stroma thylakoids). The space between the thylakoid membranes is termed the thylakoid lumen. The space outside the thylakoid membranes is termed the chloroplast stroma.

PNG rendering of a thylakoid stack. Modeled and rendered in Blender Image Source: Wikipedia Commons
Green plants possess the pigment chlorophyll with absorbance maxima in the red and blue regions of the visible light spectrum. Chlorophyll has a long phytol tail which permits anchoring of the pigment in the thylakoid membrane, and a light absorbing chromophore with Mg2+ at its center coordinated with 4 nitrogen atoms. Higher plants contain 2 chlorophylls (a and b) differing only in one substituent on the ring:
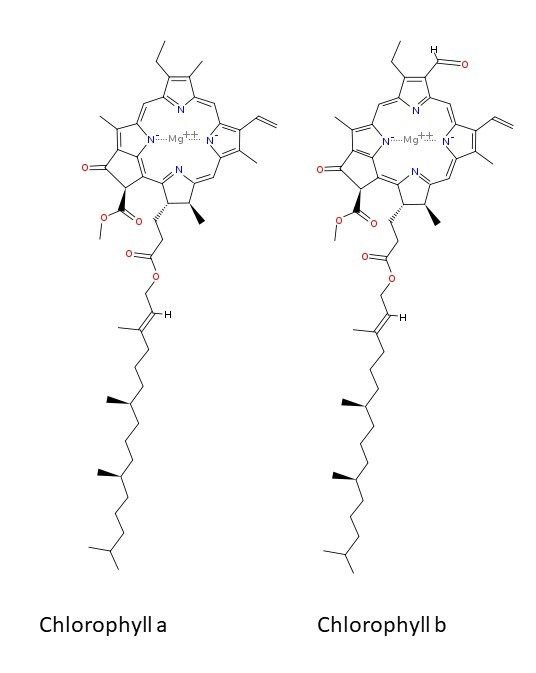
Structures of chlorophyll a and b. Image Source: Chlorophyll - Wikipedia
Photons excite electrons of the chlorophyll pigments in the chloroplasts of plant cells raising the energy level of the electrons. This excitation energy is transmitted (funneled) to a key reaction center chlorophyll of a protein complex called photosystem II, designated P680, embedded in the chloroplast thylakoid membrane. PSII is surrounded by multiple chlorophyll molecules. From the key reaction center chlorophyll (P680) an electron is ultimately lost and, together with a proton (from the chloroplast stroma), is used to reduce a lipid soluble acceptor, plastoquinone (PQ).
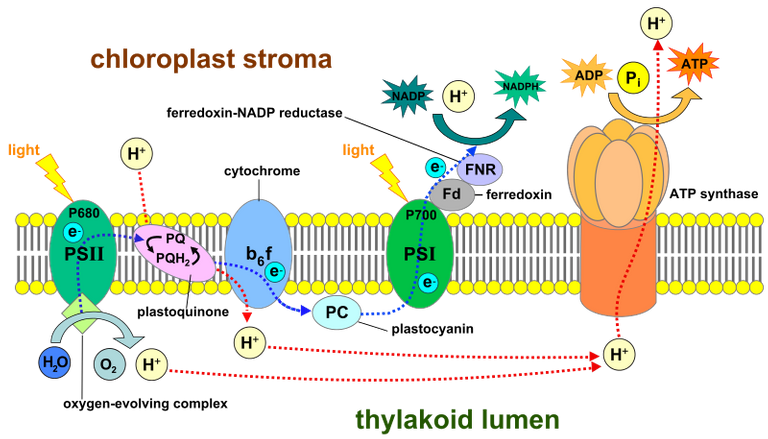
Light-dependent reactions of photosynthesis at the thylakoid membrane. Image Source: Light-dependent reactions - Wikipedia
The reaction center chlorophyll now has an electron hole which is here designated P680'. This electron hole must be filled and this electron comes from the closely associated water-splitting complex of PSII, also known as the oxygen-evolving complex. Essentially this complex catalyzes the following manganese-dependent reaction:
2H2O --> 4H+ + 4e- + O2
The protons (4H+) are dumped inside the thylakoid lumen (thereby acidifying the thylakoid lumen), oxygen gas (O2) is mostly evolved, and the 4 electrons (e-) are used to convert P680' --> P680 four times in rapid succession (P680 and P680' are continually recycled in this process) .
As noted above electrons, together with protons from the chloroplast stroma reduce plastoquinone PQ to form reduced plastoquinone (PQH2). As plastoquinone and its reduced form are lipid soluble they are free to migrate within the two lipid bilayers. The electrons of PQH2 are used to reduce a small protein on the thylakoid lumen side of the membrane, plastocyanin (PC). This reduction of plastocyanin is carried out by the cytochrome b6f complex. Upon reduction of plastocyanin further protons are dumped into the thylakoid lumen by the cytochrome b6f complex as PQ is regenerated from PQH2.
The net result of water splitting and proton pumping by the cytochrome b6f complex is acidification of the thylakoid lumen and alkalinization of the chloroplast stroma. This proton gradient is used to synthesize ATP (from ADP and phosphate (Pi)) by ATP synthase embedded in the thylakoid membrane.
Reduced plastocyanin is water soluble and mobile within the thylakoid lumen and is accessible to photosystem I (PSI). Like PSII, PSI has a key reaction center chlorophyll. In the case of PSI this reaction center chlorophyll is designated P700. Again photons excite this reaction center chlorophyll and an electron is lost and used to reduce ferredoxin (Fd) on the chloroplast stroma side of the membrane. As P700 donates an electron to ferredoxin it leaves an electron hole (P700') that is filled with an electron from the reduced form of plastocyanin. Plastocyanin returns to its oxidized form. The reduced form of ferredoxin is then used to reduce NADP+ to NADPH in the chloroplast stroma by ferredoxin:NADP+ reductase (FNR).
As we will see in the next article in this series, the chloroplast stroma houses all the enzymes of the Calvin cycle that catalyze the carbon reactions of photosynthesis using the NADPH and ATP generated by the light reactions as energy sources. The Calvin cycle enzymes have alkaline pH optima, taking advantage of the alkalinization of the stroma that takes place during the light reactions.
The ratio of NADPH and ATP produced in the light reactions can be finely tuned by a process known as cyclic electron transport. In essence this involves diversion of the reduced form of ferredoxin away from NADPH synthesis. The reduced form of ferredoxin can be used instead to reduce plastoquinone on the stroma side of the membrane thereby using energy to drive formation of a proton gradient and hence ATP synthesis. This reaction is catalyzed by ferredoxin:plastoquinone reductase.
The water-splitting complex of PSII must work in close synchrony with the light-initiated electron transport chain leading to P680' formation and reduction of PQ. Thus, electrons from water splitting are critical for regenerating P680 from P680'. Excess light (especially under environmental stress conditions such as drought, high temperature or chilling stress) can disrupt this synchrony and lead to the generation of reactive oxygen species from the water splitting complex. These molecules can damage the critical D1 protein of PSII carrying out the water-splitting reaction. Accessory pigments (yellow carotenoids) may assist in mopping up these reactive oxygen species. However, if these defenses are overwhelmed and damage to D1 occurs (termed photoinhibition), this D1 protein must be excised from the complex, degraded and a new D1 protein inserted.
PSI may still continue to function in the absence of a functional PSII if cyclic electron transport (as described above) is initiated, but this reduces photosynthesis efficiency.
This entire process is summarized in the following image:
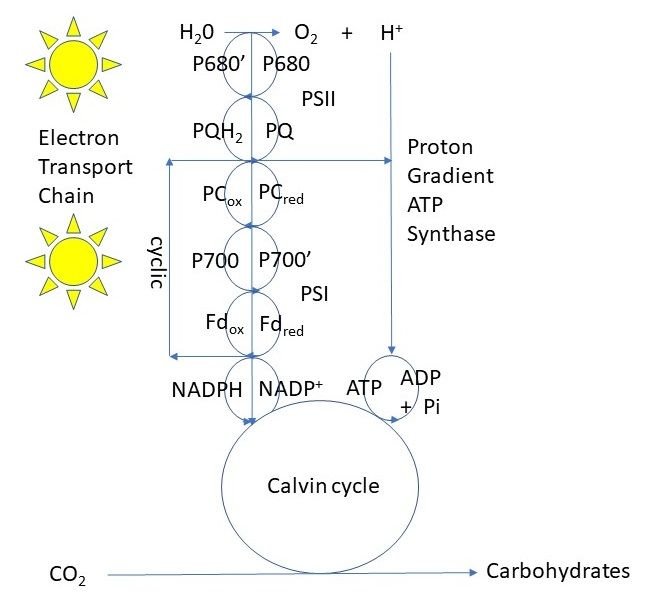
Light reactions of photosynthesis summarized (D. Rhodes)
I have simplified this scheme considerably. I have not gone into any great depth about all of the components of the electron transport chain, and the exact protein compositions (subunit structures) of the various components of the electron transport chain are omitted. For more detail please consult the following references.
References
Please feel free to leave comments or ask questions below.
Nice one @davidrhodes124
You must have heard of the artificial leafs which can do the photosynthesis to produce oxygen and it has been planned to have them almost everywhere so to resolve the future problem of air pollution.
Here is the link https://www.zmescience.com/ecology/green-living/silk-leaf-first-biological-leaf-055343/
Cheers
Excellent contribution. Thank you!
Thank you for your detailed explanation of this crucial phenomenon.
My pleasure!
Nice post @davidrhodes124! I love how the renditions of the thylakoid stacks utilize the concept of ultimate surface area to maximize production. Its amazing how much surface area plays a part in nature from small to large.
Thanks. The rendition is not mine. But it sure is quite fascinating how much surface area is achieved from this structure.
There are some places where photosynthesis can come in handy. I read sometime ago that it can be used to generate electricity (through photosynthetic algae).
Nice piece there
Is it this article to which you are referring?
Gadja et al. Self-sustainable electricity production from algae grown in a microbial fuel cell system. Biomass and Bioenergy Volume 82, November 2015, Pages 87-93
Wow! This looks more detailed than the one I was referring to. Thanks for sharing
Photosynthesis by plants is one complex topic i couldn't wrap my head around in high school. O am sure its still well wide enough like what i just read now. Thanks for the breakdown.
Thank you!